разнообразие признаков и свойств у особей и групп особей любой степени родства. И. присуща всем живым организмам, поэтому в природе отсутствуют особи, идентичные по всем признакам и свойствам. Термин «И.» употребляется также для обозначения способности живых организмов отвечать морфофизиологическими изменениями на внешние воздействия и для характеристики преобразований форм живых организмов в процессе их эволюции. И. можно классифицировать в зависимости от причин, природы и характера изменений, а также целей и методов исследования. Различают И. наследственную (генотипическую) и ненаследственную (паратипическую); индивидуальную и групповую; прерывистую (дискретную) и непрерывную; качественную и количественную; независимую И. разных признаков и коррелятивную (соотносительную); направленную (определённую, по Ч. Дарвину) и ненаправленную (неопределённую, по Ч. Дарвину); адаптивную (приспособительную) и неадаптивную. При решении общих проблем биологии и особенно эволюции наиболее существенно подразделение И., с одной стороны, на наследственную и ненаследственную, а с другой — на индивидуальную и групповую. Все категории И. могут встречаться в наследственной и ненаследственной, групповой и индивидуальной И.
Наследственная И. обусловлена возникновением разных типов мутаций (См. Мутации) и их комбинаций в последующих скрещиваниях. В каждой достаточно длительно (в ряде поколений) существующей совокупности особей спонтанно и ненаправленно возникают различные мутации, которые в дальнейшем комбинируются более или менее случайно с разными уже имеющимися в совокупности наследственными свойствами. И., обусловленную возникновением мутаций, называют мутационной, а обусловленную дальнейшим перекомбинированием генов в результате скрещивания — комбинационной. На наследственной И. основано всё разнообразие индивидуальных различий, которые включают: а) как резкие качественные различия, не связанные друг с другом переходными формами, так и чисто количественные различия, образующие непрерывные ряды, в которых близкие члены ряда могут отличаться друг от друга сколь угодно мало; б) как изменения отдельных признаков и свойств (независимая И.), так и взаимосвязанные изменения ряда признаков (коррелятивная И.); в) как изменения, имеющие приспособительное значение (адаптивная И., рис. 1), так и изменения «безразличные» или даже снижающие жизнеспособность их носителей (неадаптивная И.). Все эти типы наследственных изменений составляют материал эволюционного процесса (см. Микроэволюция). В индивидуальном развитии организма проявление наследственных признаков и свойств всегда определяется не только основными, ответственными за данные признаки и свойства генами, но и их взаимодействием со многими другими Генами, составляющими Генотип особи, а также условиями внешней среды, в которой протекает развитие организма (рис. 2 и 3).
В понятие ненаследственной И. входят те изменения признаков и свойств, которые у особей или определённых групп особей вызываются воздействием внешних факторов (питание, температура, свет, влажность и т. д.). Такие ненаследственные признаки (Модификации) в их конкретном проявлении у каждой особи не передаются по наследству, они развиваются у особей последующих поколений лишь при наличии условий, в которых они возникли. Такая И. называется также модификационной (рис. 4). Например, окраска многих насекомых при низкой температуре темнеет, при высокой — светлеет; однако их потомство будет окрашено независимо от окраски родителей в соответствии с температурой, при которой оно само развивалось (см. Морфозы, Фенокопия). Существует ещё одна форма ненаследственной И. — так называемые длительные модификации, часто встречающиеся у одноклеточных организмов, но изредка наблюдаемые и у многоклеточных. Они возникают под влиянием внешних воздействий (например, температурных или химических) и выражаются в качественных или количественных отклонениях от исходной формы, обычно постепенно затухающих при последующем размножении. Они основаны, по-видимому, на изменениях относительно стабильных цитоплазматических структур.
Между ненаследственной и наследственной И. существует тесная связь. Ненаследственных (в буквальном смысле) признаков и свойств нет, так как ненаследственные изменения являются отражением наследственно обусловленной способности организмов отвечать определёнными изменениями признаков и свойств на воздействия факторов внешней среды. При этом пределы ненаследственных изменений определяются нормой реакции (См. Норма реакции) генотипа на условия среды.
Наследственную и ненаследственную И. изучают как внутри отдельных совокупностей живых организмов, когда исследуют различия признаков отдельных особей (индивидуальная И.), так и при сравнении между собой различных совокупностей особей (групповая И.); в основе любых межгрупповых различий также лежит индивидуальная И. Даже в пределах близкородственных групп нет абсолютно идентичных особей, которые не различались бы по степени выраженности каких-либо наследственных или ненаследственных признаков и свойств. Ввиду сложности организации живых систем, даже у генотипически идентичных (например, однояйцевые близнецы) и развивающихся в практически одинаковых условиях особей всегда можно обнаружить хотя бы незначительные морфофизиологические различия, связанные с неизбежными флуктуациями условий среды и процессов индивидуального развития. Групповая И. включает различия между совокупностями любых рангов — от различий между небольшими группами особей в пределах популяции до различий между царствами живой природы (животные — растения). В сущности, вся систематика организмов построена на сравнительном анализе групповой И. Для изучения пусковых механизмов эволюционного процесса особое значение имеют различные формы внутривидовой групповой И. (см. Видообразование). Большинство видов распадается на подвиды или географические расы. В случае полной изоляции (См. Изоляция) географических форм они могут резко различаться по одному или нескольким признакам. Популяции, населяющие обширные территории и не разделённые резкими изолирующими барьерами, могут (благодаря перемешиванию и скрещиванию) постепенно переходить друг в друга, образуя количественные градиенты по тем или иным признакам (клинальная И.). Географическая, в том числе и клинальная, И. в природных условиях — результат действия изоляции, естественного отбора и др. факторов эволюции, приводящих к разделению исходной группы особей в ходе исторического формирования вида на две или несколько групп, различающихся по численным соотношениям генотипов (рис. 5). В некоторых случаях различия между группами особей в пределах вида не связаны с различиями их генотипического состава, а обусловливаются модификационной И. (различными реакциями сходных генотипов на разные внешние условия). Так называемая сезонная И. обусловлена влиянием на развитие соответствующих поколений разных погодных условий (например, у некоторых насекомых и травянистых растений, дающих два поколения в год, весенние и осенние популяции различаются рядом признаков) (рис. 6). Иногда сезонные формы могут быть результатом отбора разных генотипов (например, рано- и поздноцветущие формы трав на сенокосных лугах: в течение многих поколений устранялись особи, цветущие летом, во время сенокоса). Большой интерес представляет экологическая И. — различия между группами особей одного вида, растущими или живущими в разных местах (возвышенности и низменности, заболоченные и сухие участки и т. д.). Часто такие формы называются Экотипами. Возникновение экотипов также может быть результатом как модификационных изменений, так и отбора генотипов, лучше приспособленных к местным условиям. Наследственной И. обусловлены различные формы внутрипопуляционного Полиморфизма. В некоторых популяциях наблюдается сосуществование двух или более ясно различимых форм (например, у двухточечной божьей коровки почти во всех популяциях встречаются чёрная форма с красными пятнами и красная форма с чёрными пятнами). В основе этого явления могут лежать разные эволюционные механизмы: неодинаковая приспособленность сосуществующих форм к условиям различных сезонов года, повышенная жизнеспособность гетерозигот, в потомстве которых постоянно выщепляются обе гомозиготные формы или другие, ещё недостаточно изученные механизмы. Таким образом, и групповая, и индивидуальная И. включают изменения как наследственной, так и ненаследственной природы.
Независимой И. признаков противопоставляют коррелятивную И. — взаимосвязанное изменение различных признаков и свойств: связь между ростом и весом особей (положительная корреляция) или темпом клеточного деления и величиной клеток (отрицательная корреляция). Корреляции могут быть обусловлены чисто генетическими причинами (Плейотропия) или взаимозависимостями процессов становления определённых признаков и свойств в индивидуальном развитии особей (онтогенетические корреляции), а также сходными реакциями разных признаков и свойств на одни и те же внешние воздействия (физиологические корреляции). Наконец, корреляции могут отражать историю происхождения популяций из смеси двух или более форм, каждая из которых привносит не отдельные признаки, а комплексы взаимосвязанных признаков и свойств (исторические корреляции). Изучение коррелятивной И. имеет важное значение в палеонтологии (например, при реконструкции вымерших форм по отдельным ископаемым остаткам), в антропологии (например, при восстановлении черт лица на основе изучения черепа), в селекции и медицине.
Основные методы изучения И. — сравнительно-описательный и биометрический (см. Биометрия). Совокупность этих методов позволяет исследовать как паратипическую, так и генотипическую компоненты общей фенотипической И. Так, первую можно изучать, сравнивая генотипически идентичные Клоны и чистые линии (См. Чистая линия), развивающиеся в разных условиях. Сложнее выделить чисто генотипическую И. из общей фенотипической. Это возможно сделать на основе биометрического анализа (см. Наследуемость). В медицинской генетике для тех же целей используется определение процента конкордантности (совпадения) тех или иных признаков у одно- и разнояйцевых близнецов.
Наследственность и И. живых организмов иногда противопоставляют как «консервативное» и «прогрессивное» начала. В действительности же они теснейшим образом связаны. Отсутствие полной стабильности генотипа обусловливает мутационную и (в ходе дальнейших скрещиваний и расщеплений) комбинационную И., т. е. в целом — генотипическую И. Паратипическая (ненаследственная) И. — результат лишь относительной стабильности генотипа при определении им в онтогенезе нормы реакции при развитии признаков и свойств особей. Из этого следует возможность экспериментальных воздействий как на наследственную, так и на ненаследственную И. Первую можно усилить воздействием мутагенных факторов (излучения, температура, химические вещества). Размах и направление комбинационной И. можно контролировать с помощью искусственного отбора (См. Искусственный отбор). На ненаследственную И. можно воздействовать, изменяя условия среды (питание, свет, влажность и т. д.), в которых протекает развитие организма.
Чёткое представление о категориях и формах И. необходимо при построении эволюционных схем и теорий, так как явления наследственности и И. лежат в основе эволюционного процесса, а также в практической селекции растений и животных, при изучении ряда проблем медицинской географии и популяционной антропологии.
Лит.: Филипченко Ю. А., Изменчивость и методы её изучения, 2 изд., Л., 1926; Четвериков С. С., О некоторых моментах эволюционного процесса с точки зрения современной генетики, «Журнал экспериментальной биологии», 1926, т. 2, № 1; Иогансен В., Элементы точного учения об изменчивости и наследственности с основами вариационной статистики, М. — Л., 1933; его же, О наследовании в популяциях и чистых линиях, М. — Л., 1935; Холден Дж., Факторы эволюции, пер. с англ., М. — Л., 1935; Дарвин Ч., Происхождение видов, ..., Соч., т. 3, М., 1939; Шмальгаузен И. И., Организм, как целое в индивидуальном и историческом развитии, [2 изд.], М. — Л., 1942; Астауров Б. Л., Изменчивость, в кн.: Большая медицинская энциклопедия, т. 11, М., 1959; Вавилов Н. И., Закон гомологических рядов в наследственной изменчивости, Избр. произв., т. 1, Л., 1967, с. 7—61; его же, Линнеевский вид как система, там же, с. 62—87; Лобашев М. Е., Генетика, 2 изд., Л., 1967; Майр Э., Зоологический вид и эволюция, пер. с англ., М., 1968; Тимофеев-Ресовский Н. В., Воронцов Н. Н., Я блоков А. В., Краткий очерк теории эволюции, М., 1969; Fisher R., The genetical theory of natural selection, Oxf., 1930; Falconer D., Introduction to quantative genetics, Edinburgh — L., 1960.
Н. В. Тимофеев-Ресовский, Е. К. Гинтер, Н. В. Глотов, В. И. Иванов.
Изменчивость у микроорганизмов. У микроорганизмов, как и у других организмов, различают ненаследственную и наследственную И. Изменению могут подвергаться любые морфологические и физиологические признаки: величина и форма микроорганизмов, вид и окраска их колоний, способность усваивать или синтезировать различные органические вещества, болезнетворность и др. Наследственная И. микроорганизмов — результат мутаций, возникающих спонтанно или вызываемых физическими или химическими мутагенами (ультрафиолетовые лучи, ионизирующая радиация, этиленимин и др.). У мутантов могут резко усиливаться или снижаться такие количественные признаки, как способность к биосинтезу аминокислот, антибиотиков, ферментов, витаминов и т. п. Возникают так называемые дефицитные мутанты, способные расти только при добавлении к среде определённых аминокислот, пуринов, пиримидинов и др. Микроорганизмы размножаются очень быстро. Поэтому на них легче изучать все формы И., а также осуществлять искусственный отбор полезных мутантов (см. Селекция). Так, при непрерывном культивировании соответствующих микроорганизмов (проточные культуры) в питательной среде, содержащей, например, антибиотик, фенол или сулему, легко могут быть получены формы, устойчивые к данному веществу (адаптивная И.). Наблюдаются у микроорганизмов и взаимосвязанные изменения (коррелятивная И.). Так, возникновение у болезнетворных микробов складчатых колоний сопровождается снижением их иммуногенности. У микроорганизмов, имеющих истинный половой процесс (некоторые плесневые грибы, спорогенные дрожжи), возможно скрещивание, сопровождающееся перекомбинированием генов и получением гибридов. У несовершенных грибов и бактерий, лишённых истинного полового процесса, такие гибриды не могут быть получены.
А. А. Имшенецкий.

Рис. 2. Наследственная изменчивость форм роста у капусты: 1 — дикая однолетняя; 2 — лиственная; 3 — савойская; 4 — кормовая; 5 — брюссельская; 6 — брокколи; 7 — кольраби; 8 — цветная; 9 — кочанная.

Рис. 3. Наследственная изменчивость формы гребня у петухов: А — гороховидный; Б — розовидный; В — листовидный; Г — ореховидный.

Рис. 4. Ненаследственная изменчивость величины клеток у инфузорий: вариация размеров в каждом из последующих клонов не зависит от размера исходной особи.

Рис. 5. Географическая изменчивость формы листа у растений ветреницы из различных районов Европы.
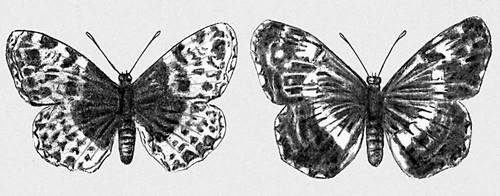
Рис. 6. Сезонная изменчивость у бабочки пестрокрыльницы; слева — весенняя форма, справа — летняя.
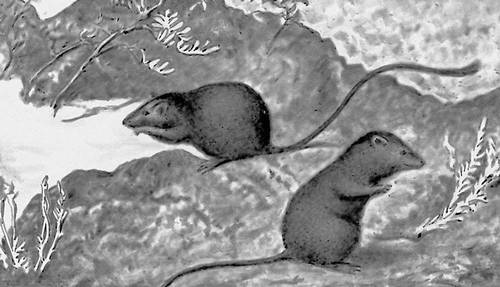
Рис. 1a. Черная адаптивная окраска у мышей Perognathus, живущих на чёрной лаве.

Рис. 1б. Белая адаптивная окраска у мышей Perognathus, живущих на песках.
Большая советская энциклопедия. — М.: Советская энциклопедия. 1969—1978.