- Семейство марантовые (Maranthaceae)
-
Семейство марантовых, обнаруживающее черты наиболее высокой организации в порядке имбирных, объединяет около 30 родов и 400 видов, распространенных главным образом в тропической зоне всех континентов, за исключением Австралии. Немногие виды марантовых произрастают в субтропиках и лишь редко встречаются в зоне умеренно теплого климата. Особенно богата марантовыми флора Южной и Центральной Америки, включающая 11 родов этого семейства и среди них самый большой — калатея (Calathea, рис. 229, 230), который объединяет около 150 видов. Наиболее широкое распространение с севера на юг в западном полушарии обнаруживают виды рода талия (Thalia). Талия беловатая (Т. dealbata) встречается в Техасе, Луизиане и юго-восточных штатах США до 37° с. ш., а талия многоцветковая (Т. multiflora) достигает в Уругвае 33° ю. ш. Талия коленчатая (Т. geniculata) растет на болотах Флориды, а к югу распространена до Аргентины; этот вид встречается и в Западной Африке. В Африке марантовые представлены главным образом эндемичными родами и преобладают во влажном тропическом климате западной части континента от Сьерра-Леоне на севере до Анголы на юге. В восточной части материка произрастают лишь немногие виды. На Мадагаскаре из марантовых встречаются лишь галопегия Перрье (Halopegia perrieri) и ктенофриниум односторонний (Ctenophrynium unilaterale), представляющий монотипный эндемичный род. В тропической Азии наибольшее число видов марантовых произрастает в лесах полуострова Малакка и на Яве, немногие виды встречаются в Индии и на острове Шри-Ланка. Среди них наиболее широкое распространение имеет фриниум головчатый (Phrynium capitatum), растущий в Индии, Бирме, Юго-Западном Китае и доходящий к югу до Явы. С этим видом по величине ареала могут соперничать лишь некоторые виды рода донакс (Donax), распространенные в Индии, Бирме, на полуостровах Индокитай и Малакка, на островах Малайского архипелага и Океании.
, Марантовые — характерные обитатели тропических дождевых лесов, где они образуют нередко непроходимые заросли. Они часто встречаются на низких болотистых и даже затопляемых местах, по берегам водоемов, вдоль рек и ручьев. Шуманниантус вильчатый (Schumannianthus dichotomus) произрастает в мангрове восточного полушария вместе с пальмой нипой или поселяется на рисовых полях. Некоторые марантовые обитают в районах с засушливым периодом в листопадных и полулистопадпых лесах, переживая засуху в виде корневищ. К таким видам относятся, например, калатея широколистная (Calathea latifolia) и калатея крупночашелистиковая (С. macrosepala) из Южной Америки; последняя особенно засухоустойчива. Марантовые являются частыми обитателями вторичных формаций, образуя сплошные заросли на вырубках или в заброшенных ирригационных каналах. Большинство видов произрастает на высоте до 1000 м над уровнем моря, рекорд высотного распространения представляет калатея Тимоти (С. timothei), растущая по склонам Анд на высоте 2100 м.
Марантовые — характерные обитатели тропических дождевых лесов, где они образуют нередко непроходимые заросли. Они часто встречаются на низких болотистых и даже затопляемых местах, по берегам водоемов, вдоль рек и ручьев. Шуманниантус вильчатый (Schumannianthus dichotomus) произрастает в мангрове восточного полушария вместе с пальмой нипой или поселяется на рисовых полях. Некоторые марантовые обитают в районах с засушливым периодом в листопадных и полулистопадпых лесах, переживая засуху в виде корневищ. К таким видам относятся, например, калатея широколистная (Calathea latifolia) и калатея крупночашелистиковая (С. macrosepala) из Южной Америки; последняя особенно засухоустойчива. Марантовые являются частыми обитателями вторичных формаций, образуя сплошные заросли на вырубках или в заброшенных ирригационных каналах. Большинство видов произрастает на высоте до 1000 м над уровнем моря, рекорд высотного распространения представляет калатея Тимоти (С. timothei), растущая по склонам Анд на высоте 2100 м.
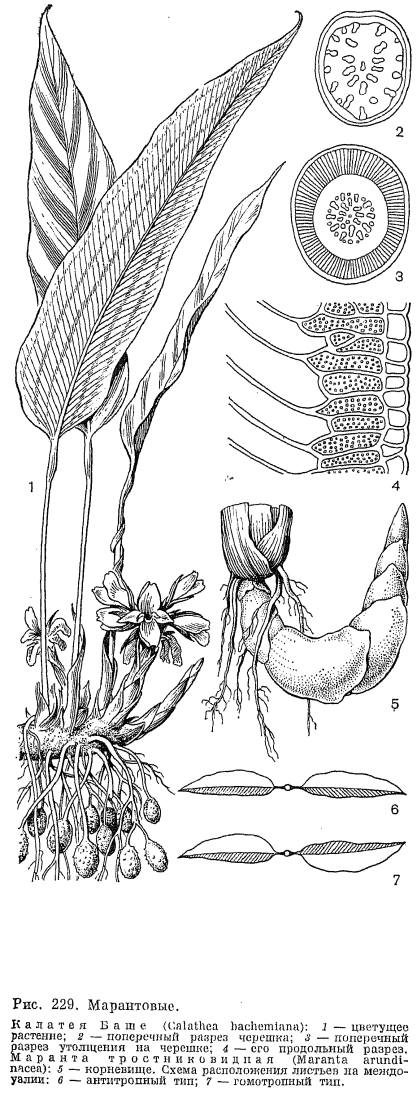
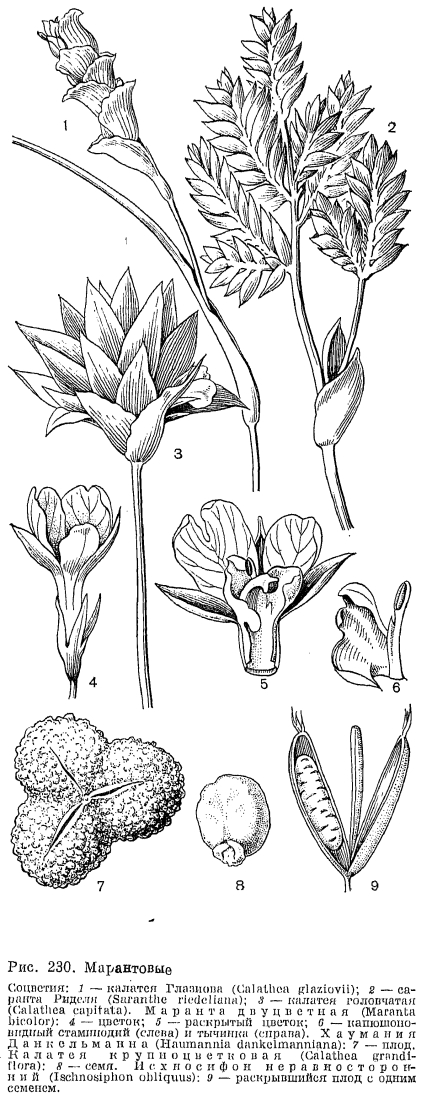
,Марантовые — многолетние травянистые растения с симподиально ветвящимися корневищами. Корневища их волокнистые и иногда жесткие, деревянистые, покрыты двурядно расположенными чешуевидными, нередко опушенными листьями. У некоторых видов корневища образуют направленные в глубь почвы утолщенные столоны, в которых запасаются вода и питательные вещества. Такие клубневидные столоны образует маранта тростникавидная (Maranta arundinacea, рис. 229, 5). Многие виды маранты, калатеи, саранты (Saranthe) и других родов имеют клубневидно утолщенные корни. У калатеи крупночашелистиковой корневые клубни достигают в диаметре 6 см. У маранты двуцветной (М. bicolor, рис. 230, 4 — 6) утолщено основание цветоносного стебля. Корневища марантовых обычно имеют короткие междоузлия, так что надземные побеги образуют густые куртины. Стебли марантовых большей частью хорошо развиты, симподиально ветвятся, иногда достигают большой высоты и становятся деревянистыми. Для марантовых характерно образование междоузлий, очень неравных по длине. Внизу стебель развивает обычно влагалищные листья на очень коротких междоузлиях, а междоузлие, несущее первый лист с развитой пластинкой, как правило, длиннее остальных. У видов донакса оно имеет длину около 3 м, а последующие междоузлия так коротки, что листья, собранные густым пучком, венчают его на вершине. Некоторые виды строманты (Stromanthe), миросмы (Myrosme) и других родов образуют над пучком сближенных листьев снова длинное междоузлие, несущее второй ярус густо собранных листьев. Виды африканских родов гипселодельфис (Hypselodelphis), трахифриниум (Trachyphrynium) и некоторые виды южноамериканского рода исхносифон (Ischnosiphon, рис. 230, 9) имеют бамбуковидный габитус. Их стебли растут быстро в высоту, сохраняя почти одинаковый диаметр, на их утолщенных узлах имеются чешуевидные листья, которые вскоре опадают. Из пазушных почек развиваются симподиально ветвящиеся боковые побеги. Для марантовых характерно наличие предлиста в основании каждой ветви. Предлист имеет два килевидных продольных выступа на стороне, прижатой при развитии к главному стеблю. У видов гипселодельфиса предлистья достигают в длину 10 см; большей частью они мелкие, опадающие. После предлиста ветвь обычно несет влагалищный лист, а затем уже листья с развитой пластинкой. Ветви у марантовых отходят под определенным для каждого вида углом, образуя зигзаг или завиток. Такой характер ветвей способствует их закреплению на ветвях соседних кустарников и деревьев. Таким образом стебли гипселодельфиса Цеикера (H. zenkerianum), растущего в тропических лесах Западной Африки, поднимаются, цепляясь ветвями за окружающие деревья, на высоту до 10 м. Ветви и оси соцветий хауманнии Данкельманна (Haumannia dankelmanniana) оснащены крючковидно загнутыми колючками, которыми растение цепляется за листья и ветви деревьев и создает таким образом непроходимые заросли в лесах Западной Африки. Многие виды калатеи, маранты, фриниума и других родов имеют, напротив, очень короткий стебель с розеткой листьев. Число листьев, развиваемых на стеблях до образования соцветий, как правило, определенное у каждого вида. У некоторых марантовых оно сокращено до одного листа, обычно вертикально стоящего на длинном черешке. Такой габитус имеют виды африканских родов мегафриниум (Megaphrynium), афрокалатея (Afrocalathea) и тауматококкус (Thaumatococcus), единственный вид которого тауматококкус Даниэля (Т. daniellii) имеет черешок листа длиной 2 — 3 м.Листья марантовых расположены двурядно на стебле и имеют хорошо развитое незамкнутое влагалище, у некоторых видов с лигулой. На черешке листа около прикрепления пластинки всегда имеется особое утолщенное сочленение (лат. pulvinus), по наличию которого марантовые легко узнать в вегетативном состоянии. Проводящие пучки, разбросанные в тканях черешка, в области этого утолщения группируются в центре, а по периферии под хлорофиллоносной палисадной тканью располагается слой длинных радиально и косо направленных тонкостенных клеток с водянистым клеточным соком (рис. 229). Колебания тургора этих клеток вызывают изменение угла наклона листа. Такой механизм позволяет регулировать положение пластинки по отношению к свету и имеет большое биологическое значение для растений, живущих в густой тени под пологом тропического леса.В течение суток листья марантовых совершают движения, улавливая солнечные лучи. Многие виды калатеи и маранты, в том числе известное комнатное растение маранта беложилковатая (М. leuconeura) из лесов Бразилии, вечером поднимают свои листья вверх и складывают их вместе, то же происходит и за несколько часов перед грозой. Таким образом эти растения предсказывают погоду. Пластинка листа марантовых овальная, продолговатая, ланцетная или почти линейная, с крупной центральной жилкой. Многочисленные боковые жилки слегка изогнуты и соединены через, определенные интервалы характерными поперечными анастомозами. Как у всех представителей порядка имбирных, листья марантовых асимметричны в результате развития внутри трубки влагалища, которая стесняет рост наружной половины трубчато свернутого листа. У некоторых марантовых узкая наружная половинка листа имеет постоянное положение справа или слева от главной жилки. Такие листья, называемые гомотропными, преобладают у калатеи, маранты, исхиосифона и других родов. У многих марантовых листья с левой узкой половинкой чередуются с листьями, у которых она расположена справа. Такие листья, называемые антитропными, характерны, например, для видов строманты и трахифриниума. Нижняя сторона листьев марантовых иногда окрашена в пурпурный или лиловатый цвет, а верхняя часто имеет пестрый рисунок за счет чередования темных и, светлых полос или в виде темных пятен на светлом фоне. Пятнистость, иногда сочетается с белыми или розовыми жилками, шелковистым блеском или бархатистой поверхностью эпидермы, клетки которой имеют прозрачные линзовидные утолщения оболочки, концентрирующие световые лучи. Темные пятна на листьях образуются за счет более высокого содержания пластид в клетках, белый цвет связан с образованием воздухоносных межклеточных пространств, а розовый и красный — с наличием антоциана в клетках. Иногда пестрая окраска присуща лишь ювенильным листьям, а взрослые растения развивают однотонные зеленые листья. Характер пестрого рисунка является систематическим признаком, позволяющим определить вид или его разновидность. Листья некоторых марантовых покрыты восковым налетом, как, например, у талии беловатой. Особенно обильный восковой налет на нижней, абаксиальной стороне имеют огромные, полутораметровой длины листья калатеи желтой (С. lutea), растущей по болотистым местам в Центральной Америке и на островах Карибского моря.Марантовые нередко имеют опушение из одноклеточных волосков с погруженным основанием. В корнях, а иногда и в стеблях и изредка в листьях марантовых имеются сосуды с простой или лестничной перфорацией.Многие марантовые зацветают в первый год жизни, но отдельные лесные виды цветут исключительно редко. Соцветия марантовых занимают верхушечное положение на облиственных стеблях или на специализированных безлистных цветоносах, вырастающих от корневища. Как правило, марантовые имеют сложные соцветия, состоящие из нескольких элементарных соцветий, которые представляют собой колосовидный тирс с двурядно расположенными кроющими листьями. У некоторых видов калатеи кроющие листья располагаются тесной спиралью, в компактных соцветиях головчатого типа. Кроющие листья зеленые или ярко окрашены, и в этом случае они, несомненно, играют роль в привлечении опылителей. Кроющие листья обычно ладьевидно сложены и содержат в пазухах парциальные соцветия, которые представляют серповидную серию последовательно развивающихся монохазиев. У калатеи в парциальном соцветии может развиваться от 4 до 13 монохазиев, а у исхносифона неравностороннего (Ischnosiphon obliquus) их число достигает 17. В то же время у южоамериканского рода монофилланта (Monophyllanthe) и некоторых видов саранты парциальное соцветие состоит лишь из одного монохазия. В основании монохазий несет обычно двухкилевой или трехкилевой предлист, два цветка монохазия зеркально-симметричны. Монохазий марантовых, по-видимому, является результатом редукции до двух цветков многоцветкового завитка, столь характерного для всего порядка имбирных. У видов южноамериканского рода строманта можно заметить рудимент третьего цветка, иногда третий цветок бывает хорошо развит. У монотагмы (Monotagma) и монофриниума (Monophrynium) произошла редукция монохазия до одного цветка, причем монотагма имеет парциальное соцветие из 3 или 4 редуцированных монохазиев, а у монофриниума в пазухе кроющего листа находится единственный цветок. В зависимости от длины осей и цветоножек тирс может быть компактным колосовидным или даже головчатого облика, а может иметь и рыхлую кистевидную структуру. У многих марантовых главная ось соцветия симподиально ветвится, благодаря чему образуются сложные соцветия с многими тирсами, венчающими каждую ветвь. В соцветиях марантовых часто имеются прицветники, обычно они пленчатые, слабо окрашенные, у многих видов они быстро опадают. Прицветнички, имеющиеся у многих марантовых, обычyо маленькие, пленчатые, а у цветков некоторых видов калатеи, фацелофриниума (Phacelophrynium) и исхносифона они нитевидные, булавовидно утолщенные на конце и, видимо, играют роль амортизаторов для нежных структур цветка, зажатых жесткими кроющими листьями. На прицветничках калатеи портобельской (С. portobelensis) имеются внецветковые нектарные желёзки, привлекающие в соцветия многочисленных муравьев.Цветки марантовых обоеполые и, так же как у канновых, асимметричные. Чашелистики обычно свободные, а лепестки у основания срастаются в трубку. Лепестки неравные по величине (адаксиальный лепесток крупнее остальных и несколько вогнут), большей частью белые или кремовые, редко розовые или голубые. Из двух кругов андроцея у марантовых полностью развит лишь внутренний. Так же как у канновых, все тычинки их имеют лепестковидное строение и срастаются с трубкой венчика у зева. Фертильна лишь одна тычинка, несущая сбоку лепестковидной нити только одно гнездо пыльника. Стаминодий, который у канновых играет роль губы, у марантовых довольно короткий, упругий, с двумя утолщенными участками, за что получил название мозолистого стаминодия. Другой стаминодий внутреннего круга имеет капюшонообразное строение и в бутоне охватывает столбик и прижатую к нему тычинку. Один край его имеет крючковидный вырост, играющий важную роль в специализированном механизме опыления марантовых. В наружном круге андроцея марантовых всегда отсутствует передний, абаксиальный член и развиты обычно один или два лепестковидных стаминодия. В цветках тауматококкуса, некоторых видов калатеи, строманты и других родов стаминодии наружного круга не развиты совсем.Единственная половина пыльника марантовых вскрывается продольно еще в бутоне. Пыльцевые зерна гладкие, с безапертурной оболочкой. Гинецей из 3 плодолистиков, синкарпный; завязь нижняя, 3- или 1-гнездная за счет недоразвития двух гнезд. В каждом гнезде почти базально расположен единственный анатропный или кампилотропный семязачаток. В тканях завязи над перегородками всегда имеются крупные овальные нектарные желёзки, открывающиеся у основания столбика. Нектар накапливается в нижней части трубки венчика, которая прикрыта иногда волосками опушения. Столбик короткий, довольно толстый, изогнут на вершине так, что рыльце, находящееся в воронковидном углублении, направлено несколько вниз и в сторону от тычинки, и пыльца своего пыльника на него не попадает, а высыпается в особое углубление над рыльцем. Клейкие выделения клеток, выстилающих края углубления, не дают пыльце высыпаться в полость бутона. В бутоне лепестки спирально свернуты, а крючковидный столбик приподнят, несколько выпрямлен краем капюшоновидного стаминодия и находится в напряжении.Цветки марантовых опыляют мелкие виды пчел. Американский ученый Элен Кеннеди (1978), изучающая марантовые в природе, наблюдала на их цветках пчел из родов эвглосса (Euglossa) и эвлема (Eulaema). Пчела обычно садится на стаминодии наружного круга и по мозолистому стаминодию, как по желобу, вводит свой хоботок в цветочную трубку. На пути насекомого, пробирающегося за нектаром, словно взведенный курок, стоит крючковидный вырост капюшоновидного стаминодия. Толчок по этому триггерному устройству отодвигает этот стаминодий, приподнятый столбик освобождается и, сгибаясь, хлопает рыльцем: по насекомому, забирает имеющуюся на нем пыльцу, принесенную с других цветков, и одновременно стряхивает на него свою пыльцу. По К. Фегри и Л. Ван дер Пэйлу (1906), в цветках калатеи, исхносифона и монотагмы пыльца высыпается точно в ямку у основания хоботка, где пчела ее не достает, когда чистится. Виды пчел с длинными хоботками, а также колибри иногда «грабят» нектар в цветках калатеи, не трогая крючковидного выроста и не приводя в действие механизм опыления. Э. Кеннеди удалось проследить узкоспециализированные связи видов калатеи с видами опылителей. Например, крупные цветки калатеи Донелл — Смита (Calathea donell — smithii) опыляет пчела эвлема мериана (Eulaema meriana), а в более мелких цветках с короткой трубкой она пьет нектар, не опыляя их. Среди видов калатеи подрода псевдофриниум (Pseudophrynium) Э. Кеннеди обнаружила довольно обширную группу с неоткрывающимися цветками, в которых, однако, не происходит самоопыления. Их насильственно открывают и опыляют пчелы эвлемы. Пчела жует копчик бутона, чтобы разъединить лепестки, а затем головкой поднимает медианный лепесток и пробирается в цветок. Немногие виды калатеи имеют настоящие клейстогамные цветки, к таким относится калатея панамская (Calathea panamensis). Несколько видов калатеи имеют красные кроющие листья соцветий и желтые цветки, например калатея боковая (С. lateralis), растущая в Андах Колумбии и Перу. Сочетание красного цвета с желтым характерно для птицеопыляемых соцветий геликонии и некоторых бромелиевых. Э. Кеннеди (1977) высказывает предположение о возможном переходе к опылению птицами у калатеи. Крючковидный вырост на капюшоновидном стаминодии у этих видов редуцирован, и обычный для марантовых механизм опыления действовать не может.Плоды марантовых — трехгнездные или одногнездные локулицидные коробочки или односемянные орешковидные или ягодообразные. У видов галопегии два гнезда завязи остаются стерильными и оболочка единственного семени полностью срастается с оболочкой плода, образуя семянку. Виды саркофриниума (Sarcophrynium) и тауматококкуса Даниэля, растущие в тропической Африке, имеют сочные плоды с желеобразным эндокарпием, у последнего — очень сладкие, их охотно поедают обезьяны и другие животные. Плоды донакса канновидного (Donax cannaeformis), обитающего по морским побережьям Малайзии, Малайского архипелага и Новой Гвинеи, легко плавают в воде благодаря пробкообразному эндокарпию и разносятся течениями.Семена марантовых большей частью имеют ариллус, микропиле на семени закрыто круглой крышечкой, которую выталкивает корень зародыша при прорастании. Зародыш сильно изогнут и окружен обильным периспермом. Ариллус, несомненно, играет роль в раскрытии коробочек. У некоторых видов калатеи, стахифриниума (Stachyphrynium) и других родов ариллус имеет выросты, которые в сухом плоде сложены компактно, а набухая, распрямляются и, словно пружины, раскрывают плод. У марантовых с нераскрывающимися плодами семена обычно без ариллуса. Ариллус имеет биологическое значение и для распространения семян. Полевые наблюдения и эксперименты американских биологов К. Хорвица и А. Битти (1980), проведенные в тропических лесах на юге Мексики, убедительно доказывают участие муравьев, питающихся ариллусами калатеи, в распространении ее семян. Немалую роль в распространении семян марантовых играют небольшие птицы, живущие в подлесках влажных тропических лесов. Семена видов талии, живущих по берегам водоемов, нередко поедают утки. Семена марантовых обладают способностью покоиться, сохраняя жизнеспособность. По наблюдениям Э. Кеннеди (1978), семена видов калатеи, произрастающих в районах с засушливым периодом, не прорастают до начала новых дождей.Семейство марантовых делится на две трибы: фриниевых (Phrynieae) и марантовых (Маrаnteae). Для фриниевых характерна 3-гнездная завязь, но у видов фриниума не развит иногда один семязачаток, а у фриниума мелкоцветкового и видов галопегии недоразвитыми остаются два гнезда завязи. Триба марантовых характеризуется развитием лишь одного — переднего, абаксиального гнезда завязи.К семейству марантовых принадлежит одно из древних культурных растений — маранта тростниковидная, или вест-индский аррорут (Maranta arundinacea, рис. 229, 5). Ее веретеновидно утолщенные корневища используются как источник крахмала, который является ценным диетическим продуктом. Маранта тростниковидная произрастает в тропических лесах Южной Америки и культивируется повсюду в тропиках. Как заменитель аррорута в Гвинее выращивают маранту арума (М. аroumа). Стебли видов донакса и некоторых других марантовых расщепляют и используют для плетения корзин и других изделий. Пестролистные виды калатеи и маранты выращивают как декоративные растения в оранжереях и комнатах.
Жизнь растений: в 6-ти томах. — М.: Просвещение. Под редакцией А. Л. Тахтаджяна, главный редактор чл.-кор. АН СССР, проф. А.А. Федоров. 1974.
.