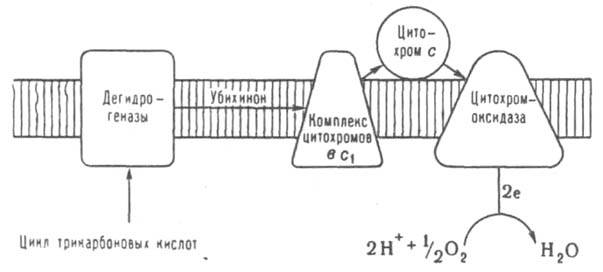
Дыхат. цепь митохондрий. Схематически изображен фрагмент митохондриальной мембраны в разрезе. Заштрихован фосфолипидный бислой. Стрелками обозначен путь электронов от субстратов окисления к О 2. Цитохромы b, с и с 1 белки-переносчики электронов; в качестве простетич. группы содержат гем. Др. важная ф-ция клеточного Д. - окислит. биосинтез большого числа нужных организму в-в. Так, напр., образование ненасыщ. жирных к-т из насыщенных, ключевые этапы синтеза простагландинов, стероидных и нек-рых пептидных гормонов, достройка поперечных сшивок между цепями коллагена в соединит. ткани идут в организме с потреблением О 2. Высокая окислит. способность О 2 используется в клеточном Д. также для разрушения и детоксикации чужеродных вредных в-в и для деградации мн. подлежащих удалению продуктов собств. метаболизма (напр., окислит. распад аминокислот, пуриновых оснований). Особую роль в детоксикации гидрофобных орг. соед. играет электронотранспортная цепь микросом, представляющих собой фракцию мембранных пузырьков, к-рую получают при дифференц. центрифугировании клеточных гомогенатов; содержит фрагменты мембран эндоплазматич. сети, комплекса Гольджи и др. Ключевой компонент микросомальной системы детоксикации цитохром Р-450 (подобно монооксигеназам он катализирует р-цию
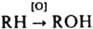 , напр., гидроксилирование стероидов; второй атом О в молекуле О 2 восстанавливается при этом до Н 2 О). Эта электронотранспортная цепь особенно активна в печени животных. В биохимии клеточного Д. различают неск. осн. р-ций с участием О 2: 1) катализируемое оксидазами ("аэробными гидрогеназами") четырехэлектронное восстановление О 2 до Н 2 О или двухэлектронное до Н 2 О 2:
, напр., гидроксилирование стероидов; второй атом О в молекуле О 2 восстанавливается при этом до Н 2 О). Эта электронотранспортная цепь особенно активна в печени животных. В биохимии клеточного Д. различают неск. осн. р-ций с участием О 2: 1) катализируемое оксидазами ("аэробными гидрогеназами") четырехэлектронное восстановление О 2 до Н 2 О или двухэлектронное до Н 2 О 2: 2RH2 +O2 : 2R + 2Н 2O; RH2 + O2 : R + H2O2
2) Включение обоих атомов О 2 в молекулу окисляемого в-ва, катализируемое диоксигеназами (оксигеназами):
RH2 + O2 : R(OH)2
3) Включение одного из атомов О 2 в молекулу окисляемого в-ва, др. атом О восстанавливается с образованием Н 2 О в результате окисления второго субстрата:
RH + R' Н 2 + О 2 : ROH + R' + Н 2 О
Ферменты, катализирующие эту р-цию, - монооксигеназы. В состав активных центров ферментов, взаимодействующих с О 2, обычно входят ионы переходных металлов (медь, гемовое или негемовое железо) или флавины (коферментные формы витамина рибофлавина). Интенсивность Д. организмов, тканей, клеток принято выражать в кол-ве О 2, потребляемого за единицу времени на единицу массы (напр., в мг О 2.мин -1 г -1). Важный показатель интенсивности Д. высших позвоночных - кол-во воздуха, вентилируемого легкими в 1 мин (наз. минутным объемом дыхания, или МОД). Эти величины служат важнейшим показателем уровня энергетич. обмена организма. У человека МОД в состоянии покоя составляет 5-8 л/мин, во время физич. работы - до 100 и более л/мин. Соед., подавляющие Д. (дыхат. яды), выключают энергообеспечение организма и потому являются быстродействующими ядами. Классич. дыхат. яды (цианиды, изоцианиды, сульфиды, азиды, СО и NO) угнетают концевой фермент дыхат. цепи митохондрий ( цитохром-с-оксидазу). Эти же соед. угнетают транспорт О 2 по организму, связываясь с гемоглобином. Др. важный класс дыхат. ядов - гидрофобные орг. в-ва, часто хиноидной природы, выступающие как антагонисты убихинона (замещенного 1,4-бензохинона), играющего ключевую роль во мн. стадиях переноса электронов по дыхат. цепи. Сильнейшие яды этого класса - токсич. антибиотики (ротенон, пирицидин, антимицин, миксотиазол), 2-гептил-4-гидроксихинолин-N-оксид; их используют в исследованиях тканевого Д. Способность к умеренному подавлению убихинон-зависимых р-ций в дыхат. цепи свойственна мн. лек. ср-вам (напр., барбитуратам), фунгицидам и пестицидам. Лит. Рэкер Э., Биоэнергетические механизмы: новые взгляды, пер. с англ., М.. 1979; Мецлер Д., Биохимия, пер. с англ.. т 2. М.. 1980. с. 361 445: Константинов А. С., Общая гидробиология, 4 изд., М., 1986. гл. 6; Скулачев В. П., Энергетика биологических мембран. М.. 1988; Molecular mechanisms of oxygen activation, N.Y., 1974; Wikstrdm M., Saraste M., The mitochondrial respiratory chain, в сб. Bioenergetics. Amst, 1984. p. 49-94. А. А. Константинов.
Химическая энциклопедия. — М.: Советская энциклопедия. Под ред. И. Л. Кнунянца. 1988.