PARASITISME
Dans la biosphère, deux types de transferts d’énergie entre organismes vivants sont dominants: la prédation , qui met en jeu des systèmes proie-prédateur, et le parasitisme , qui met en jeu des systèmes hôte-parasite. Dans les deux cas, un organisme se nourrit aux dépens d’un autre.
Il est généralement admis que la caractéristique du parasitisme est qu’il existe, en plus de la relation nourriture, une relation habitat: pour un parasite, l’hôte représente non seulement la table mais aussi la maison. De plus, le transfert d’énergie implique la mort de la proie au moment de l’acte de prédation tandis que le parasite ne tue jamais d’emblée l’individu hôte.
Il faut reconnaître que certaines situations se laissent difficilement classer. Par exemple, les ciliés de la panse des ruminants, les insectes inféodés à une plante hôte, les hématophages tels que les moustiques, les crustacés cirripèdes qui vivent enfoncés dans la peau des baleines, le perroquet Nestor notabilis qui se nourrit de lambeaux de chair arrachés à des moutons vivants sont-ils des prédateurs ou des parasites? Certains cas considérés comme inquilinisme, commensalisme ou symbiose pourraient être suspectés de parasitisme si on les examinait en détail.
Les insectes entomophages qui pondent dans le corps d’un autre insecte qu’ils ont préalablement immobilisé, et qui sera dévoré, donc tué, par les larves, sont appelés parasitoïdes , ce qui traduit l’hésitation à les considérer comme des parasites authentiques.
Enfin, certains organismes, qui strictement parlant ne diffèrent en rien des parasites, sont pour des raisons historiques et de méthodologie étudiés dans des disciplines différentes. C’est le cas des bactéries (bactériologie), des virus (virologie) et des champignons (mycologie).
1. Animaux parasites
L’hôte représentant le milieu où vit le parasite, cela implique des rapports de taille: le parasite doit être plus petit (en volume) que son hôte. Les animaux volumineux sont rarement parasites mais par contre sont souvent les plus parasités. Certains embranchements (spongiaires, échinodermes, urochordés) ne comptent pas de formes parasites, chez d’autres (cœlentérés, mollusques, vertébrés), elles sont rares ou exceptionnelles. Ailleurs, les parasites sont plus abondants mais limités à certaines classes ou familles. Parmi les crustacés, les cirripèdes, isopodes et copépodes comptent beaucoup d’espèces parasites. Dans d’autres embranchements, des classes ou des ordres entiers sont composés de parasites, opalines ou trypanosomes parmi les protozoaires flagellés; monogènes, cestodes et trématodes parmi les plathelminthes. Enfin, il existe des embranchements entièrement formés de parasites: apicomplexa (sporozoaires) chez les protozoaires, acanthocéphales chez les métazoaires.
L’hyperparasitisme est le cas où un parasite est l’hôte d’un autre parasite: amibes parasites d’opalines, haplosporidies parasites de trématodes larvaires, hyménoptères parasitoïdes d’hyménoptères, eux-mêmes parasitoïdes de lépidoptères.
L’adaptation au parasitisme
Selon la localisation du parasite chez l’hôte, on peut séparer trois types de parasitisme:
– Les ectoparasites vivent sur les téguments de l’hôte et restent ainsi en contact direct avec le milieu extérieur.
– Les mésoparasites se rencontrent dans l’hôte mais dans des organes ouverts sur le milieu extérieur (intestin, appareil urinaire ou génital par exemple).
– Les endoparasites vivent dans les tissus (musculaires, nerveux), dans des cavités fermées (hémocœle des insectes, système circulatoire des vertébrés) ou dans des cellules. Ils doivent pénétrer et sortir de l’hôte par effraction.
Le parasitisme résulte de l’adaptation d’organismes au milieu particulier qu’est le milieu vivant. Cette adaptation des parasites revêt parfois des aspects extraordinaires.
La morpho-anatomie est caractérisée par la tendance générale à une augmentation de la taille des parasites (lorsqu’on peut la comparer à celle des formes libres apparentées). Il semble exister une correspondance entre la taille du parasite et celle de l’hôte. Le plus grand nématode Placentonema gigantissima (8,5 m de longueur)) a été récolté dans le placenta des baleines. Les adaptations les plus importantes portent sur le développement et la modification d’organes (ventouses, crochets) destinés à assurer l’attachement du parasite à l’hôte. Elles sont particulièrement marquées chez les ectoparasites. On note aussi la réduction ou la disparition d’organes (locomoteurs, sensoriels) liés à la vie libre.
Les modifications sont parfois telles qu’il est difficile de situer le parasite dans la systématique. La sacculine adulte, simple sac appendu sous l’abdomen des crabes, ne ressemble en rien à un crustacé. Seuls les stades libres (Nauplius et Cypris ) qui assurent la dispersion de la sacculine permettent, par leur morphologie peu modifiée, de les rapprocher des crustacés cirripèdes (cf. ONTOGENÈSE ANIMALE, fig. 1).
Une des principales adaptations anatomiques est l’augmentation, parfois extraordinaire, de l’appareil reproducteur. D’autres sont liées à l’alimentation: augmentation du volume du système digestif chez les hématophages (sangsues, tiques) mais disparition chez certains parasites intestinaux (cestodes, acanthocéphales).
La physiologie des parasites peut être très voisine de celle des animaux libres chez les ectoparasites, alors qu’elle se modifie chez les mésoparasites et les endoparasites. Ainsi, en ce qui concerne la nutrition, l’absorption transtégumentaire prend souvent le pas sur la digestion de matières ingérées; par des mécanismes de diffusion simple, de transfert facilité, de transport actif et même d’endocytose au niveau des membranes plasmiques périphériques, elle permet l’acquisition de molécules très variées aux dépens du milieu hôte environnant. Il est curieux de noter que, dans un même habitat, par exemple l’intestin de l’hôte, les nématodes prennent leurs nutriments par voie digestive, les cestodes et les acanthocéphales par voie transtégumentaire, tandis que les trématodes utilisent les deux voies. Certains œufs même se nourrissent par absorption à travers la coque: l’embryon des schistosomes n’évolue en miracidium que si le milieu hôte lui fournit une série de molécules bien déterminées.
Les parasites tirent l’essentiel de leur énergie de la dégradation du glucose, le plus souvent stocké sous forme de glycogène. Cette glycolyse se fait suivant des voies comparables à celles des animaux libres, mais les processus anaérobies y jouent un rôle plus important. Les parasites ne négligent cependant pas la possibilité, quand elle existe, d’utiliser l’oxygène, même rare; certains d’entre eux (nématodes) possèdent des pigments respiratoires très performants, du type hémoglobine.
La reproduction sexuée se caractérise par une fertilité en général très importante. Elle est parfois accompagnée d’une multiplication asexuée rapide et peu consommatrice d’énergie (schizogonie, sporogonie des protozoaires, polyembryonie des hyménoptères parasitoïdes, multiplication larvaire des trématodes dans le mollusque hôte intermédiaire).
L’évitement immunitaire
Le milieu parasité est vivant, donc capable de se défendre. Qu’il s’agisse d’un invertébré, où les mécanismes de défense sont relativement simples et peu spécifiques, ou d’un vertébré, où l’immunité atteint un haut degré de complexité et de spécificité, on est étonné de voir l’hôte «accepter» le parasite.
L’évitement immunitaire se réalise par des mécanismes divers. L’un des plus étonnants (variation antigénique) est utilisé par certains trypanosomes (protozoaires, agents de la maladie du sommeil, transmis par la mouche tsé-tsé). La maladie est caractérisée par des «vagues» de forte parasitémie séparées par des «creux» où le parasite est quasi indétectable; l’intervalle est de 5 ou 6 jours. La membrane plasmique du trypanosome porte une seule protéine (glycoprotéine d’un PM de 65 000 daltons, constituée de 500 acides aminés) et n’expose donc qu’un unique antigène au système immunitaire de l’hôte. Celui-ci fabrique des anticorps dirigés contre cette protéine et l’effectif des trypanosomes s’effondre au bout de quelques jours. Alors apparaît une nouvelle vague, dont tous les individus portent une même et nouvelle glycoprotéine, non reconnue par les anticorps dirigés contre celle de la première vague. Et ainsi de suite... Les trypanosomes possèdent un millier de gènes de glycoprotéines, susceptibles d’être exprimés l’un après l’autre au cours des vagues successives. Il n’existe qu’un seul site d’expression de ces gènes, ce qui explique qu’une seule glycoprotéine puisse être présente sur la membrane.
Les plasmodiums, agents des paludismes, ne sont exposés que pendant des temps très brefs au système immunitaire puisque, la plupart du temps, ils sont intracellulaires. Libres dans le sang, les parasites n’expriment à leur surface qu’une seule protéine formant une couche qui se détache et se régénère sans cesse dès qu’elle est agressée par des anticorps. Le système immunitaire de l’hôte est ainsi trompé, n’attaquant qu’une protéine que le parasite renouvelle aisément.
Quant aux schistosomes, agents des bilharzioses, ils se recouvrent, dès leur pénétration, de molécules empruntées à l’hôte: ainsi déguisé, le parasite n’est plus reconnu comme non-soi.
Dans la plupart des cas, les parasites exercent aussi un effet immunodépresseur dû à la libération de substances particulières. Certains insectes parasitoïdes injectent dans le corps de leur victime leurs œufs porteurs d’un virus (baculovirus), et c’est ce dernier qui provoque l’immunodépression.
Lorsque l’évitement immunitaire est parfait, parasite et hôte constituent un système au sens complet du terme: il arrive que le développement d’un trématode chez un mollusque ne s’accompagne ni d’une réaction ni d’une pathologie. Simplement, le système hôte-parasite, au lieu d’affecter ses ressources à la production des œufs de l’hôte, les affecte à la production des larves du parasite. Il existe cependant des cas de pathologie parasitaire aboutissant à la mort de l’hôte (parasites se développant ou s’accumulant dans un organe vital). Lorsque la pathogénie intéresse seulement les gonades, elles peut se traduire par une castration parasitaire susceptible d’influer sur la démographie de la population hôte: exemple des crabes parasités par les sacculines (cf. ONTOGENÈSE ANIMALE, fig. 1).
Spécificité
De la formation des systèmes hôtes-parasites découle le concept de spécificité parasitaire. La spécificité est un mécanisme d’isolement, ce qui a parfois amené à comparer individu, population ou espèce d’hôte à des îles. Le nombre d’hôtes d’une espèce parasite permet de distinguer les spécificités:
– oioxène : une espèce parasite ne peut vivre que chez une seule espèce hôte;
– sténoxène : une espèce peut parasiter quelques espèces hôtes proches sur le plan de la systématique (même genre, par exemple);
– euryxène : une espèce parasite peut se rencontrer chez des hôtes non apparentés mais ayant entre eux une ressemblance écologique.
Il n’existe pas de liaison entre un type de spécificité et tel ou tel groupe de parasites.
La spécificité au niveau de l’espèce se reflète souvent à des niveaux systématiques plus élevés. Ainsi, les grégarines stylocephalidae ne parasitent que les coléoptères tenebrionidae, les cestodes tetrarhynchidea adultes ne se rencontrent que chez les sélaciens.
Dans les cycles hétéroxènes, la spécificité parasitaire peut être différente suivant le niveau du cycle. Chez les trématodes, la spécificité est le plus souvent oioxène vis-à-vis du mollusque premier hôte intermédiaire, euryxène pour l’invertébré deuxième hôte intermédiaire et sténoxène pour le vertébré hôte définitif.
Une des conséquences de la spécificité est de réduire les chances de compétition entre parasites d’espèces différentes. Quand, malgré tout, plusieurs espèces coexistent chez un même hôte, il y a séparation des niches, chacune se localisant sur ou dans un organe différent ou même dans une partie d’organe. Ainsi cinq Eimeria (coccidies) mésoparasites du lapin s’échelonnent-elles régulièrement le long de l’intestin (fig. 1). De plus, celles qui se rencontrent dans la même portion ne parasitent pas les mêmes cellules de la villosité intestinale et leur localisation à l’intérieur de la cellule n’est pas identique.
Cycles biologiques
La consommation d’énergie pour la recherche de la nourriture, importante pour les prédateurs, est extrêmement réduite pour les parasites; par contre, les modalités de la reproduction sont complexes. L’individu hôte étant mortel, le parasite doit le quitter pour atteindre un nouvel individu hôte, ce qui implique un passage dans le milieu extérieur. Sauf rares exceptions, le cycle biologique d’un parasite comprend une alternance de phases libres et de phases parasites.
Dans le milieu extérieur, le parasite se développe jusqu’à une forme infestante qui doit parvenir à l’hôte définitif (par convention, celui où se déroule la reproduction sexuée du parasite).
Dans le cycle d’un parasite, trois points sont essentiels: sortie de l’hôte, passage dans le milieu extérieur, arrivée et entrée dans l’hôte. Ces événements n’ont pas la même importance selon le type de parasitisme.
Les ectoparasites sont en contact direct avec le milieu extérieur; la sortie et le passage hors de l’hôte ne constituent pas un véritable changement de milieu.
Les monogènes, qui parasitent la peau ou les branchies de poissons, quittent leur hôte sous la forme d’œuf d’où sort une larve ciliée. Celle-ci nage à la recherche d’un poisson hôte, s’y accroche et gagne son habitat définitif où elle se développe en adulte. Le processus est comparable chez les crustacés et les mollusques parasites. Chez la majorité des ectoparasites d’animaux terrestres (tiques, par exemple), il existe aussi un stade libre qui assure le passage dans le milieu extérieur et l’infestation d’un nouvel hôte. La presque totalité des ectoparasites présente donc un cycle holoxène. En général l’infestation est active, les formes de dispersion (qui sont aussi infestantes) sont bien équipées pour la recherche et l’accrochage sur l’hôte.
Les mésoparasites vivent dans des habitats (intestin, etc.) ouverts sur le milieu extérieur. La sortie hors de l’hôte est aisée, mais implique un changement de milieu. Dans ce milieu différent, les parasites doivent être protégés pour poursuivre leur développement jusqu’au stade infestant. L’arrivée dans l’hôte se fait en général par voie buccale avec la nourriture. Si le parasite parvient dans le milieu extérieur sous une forme de dispersion protégée (œuf ou kyste) et si cette forme est directement infestante, le cycle est holoxène. Si cette forme doit passer par un hôte intermédiaire pour atteindre le stade infestant, le cycle est hétéroxène.
Le premier cas (cycle holoxène) domine chez les protozoaires intestinaux (flagellés, amibes, sporozoaires, ciliés). Les parasites enkystés sont évacués avec les excréments. L’infestation d’un nouvel hôte résulte de la consommation d’aliments souillés porteurs de kystes. Parmi les métazoaires, un cycle holoxène existe chez les nématodes. Leurs œufs, très résistants dans le milieu extérieur, peuvent être directement infestants ou éclore et donner une larve infestante elle aussi protégée.
Les cycles hétéroxènes sont communs chez les métazoaires parasites. La forme de dispersion, nageante dans le milieu aquatique ou protégée dans le milieu terrestre, parvient dans un hôte intermédiaire où elle devient stade infestant. Cet hôte intermédiaire appartient à une chaîne alimentaire de l’hôte définitif. La plupart des cycles des cestodes, des acanthocéphales, de beaucoup de nématodes répondent à ce schéma.
Le cycle des trématodes est plus compliqué car il comprend deux formes de dispersion séparées par une multiplication larvaire dans un mollusque hôte intermédiaire (fig. 2).
Les endoparasites ont un cycle le plus souvent hétéroxène. Sortie et entrée dans l’hôte se font par effraction. Celle-ci est le fait soit du parasite lui-même, soit d’un vecteur.
Les femelles gravides de la filaire de Médine (Dracunculus medinensis ) percent la peau de l’hôte pour libérer les larves dans l’eau. La larve infestante qui se forme dans un copépode hôte intermédiaire, ingérée par l’homme avec l’eau de boisson, traverse la paroi intestinale et donne l’adulte dans le tissu conjonctif sous-cutané.
Plus généralement, le vecteur est un invertébré hématophage (moustique, punaise, tique, sangsue etc.). Le parasite parvenu dans cet hôte avec le sang ingéré se transforme en stade infestant qui revient dans l’espèce hôte initiale à l’occasion d’un nouveau repas sanguin. Un tel cycle hétéroxène existe chez les trypanosomes, les plasmodiums, les filaires.
Dans tous les types, il peut apparaître des hôtes supplémentaires non obligatoires mais facilitant, par leur écologie, le succès du cycle. Ces hôtes, dits paraténiques sont porteurs de la forme infestante et s’intègrent dans une chaîne alimentaire de l’hôte définitif – cas du nématode Syngamus et de nombreux acanthocéphales (fig. 3).
La transmission peut prendre une allure extraordinaire. En se multipliant dans une tique, les Babesia envahissent les ovules et par là les larves de cette tique. Ces larves, dès leur naissance, peuvent transmettre la Babesia au vertébré hôte (transmission transovarienne). La larve infestante du nématode Uncinaria lucasi pénètre chez un phoque femelle et atteint la glande mammaire. Avec le lait, elle est ingérée par le jeune phoque chez qui elle donne un adulte intestinal.
La découverte des hôtes
L’existence d’une spécificité étroite, et par conséquent le peu de choix laissé aux parasites, rend la découverte des hôtes problématique, du moins en apparence. La solution la plus communément invoquée est la très grande fécondité des parasites: la découvertes des hôtes est facilitée par la simple augmentation des probabilités de rencontre, elle-même liée au grand nombre. Cependant, les organismes parasites ne comptent pas sur le seul hasard pour rencontrer leurs cibles vivantes. D’autres adaptations groupées sous le nom de mécanismes de favorisation interviennent dans beaucoup de transmissions parasitaires. Les helminthes en offrent des exemples frappants.
Le cycle des schistosomes comporte deux hôtes: un mollusque et un vertébré; du mollusque aquatique sortent des milliers de cercaires qui infestent le vertébré par pénétration transcutanée. Ainsi, Schistosoma mansoni , agent de la bilharziose intestinale, infeste l’homme au cours des baignades, des travaux ménagers de lavage et des travaux agricoles. Les cercaires, qui sont incapables de se nourrir au cours de leur nage, n’ont que quelques heures pour trouver un hôte (fig. 4). La rencontre avec celui-ci est favorisée par un rythme circadien de la sortie des cercaires qui se produit entre 9 h et 17 h, avec un maximum vers 11 h, coïncidant avec la fréquentation humaine de l’eau. Il s’agit d’une adaptation chronobiologique. On en observe d’aussi spectaculaires chez les filaires (nématodes); ainsi les microfilaires de Wuchereria bancrofti , qui évoluent chez les moustiques, se rassemblent dans les vaisseaux sanguins superficiels des malades, là ou le moustique pourra les ingérer, en fin de journée, c’est-à-dire au moment où les moustiques piquent.
Le cycle du trématode Microphallus papillorobustus comporte trois hôtes: un mollusque, un petit crustacé aquatique (gammare) et un oiseau (goéland). Du mollusque sortent les cercaires qui pénètrent chez les gammares où elles deviennent des métacercaires; l’oiseau s’infeste en avalant des gammares porteurs de métacercaires. On a montré que les métacercaires s’installent dans le système nerveux des gammares et altèrent le comportement de ces petits crustacés. Les gammares parasités restent en surface et s’agitent quand un oiseau s’approche, au lieu de s’enfouir dans la végétation et de s’immobiliser comme le font des gammares sains. On devine que ce comportement aberrant augmente fortement la probabilité de rencontre entre le trématode et l’oiseau.
Cet exemple illustre l’utilisation d’un maillon de chaîne alimentaire. Le parasite se fixe sur une proie naturelle du prédateur hôte ou la pénètre et exerce un effet pathogène de manière à l’affaiblir et à faciliter sa capture. Si le cas précédent est spectaculaire, des exemples comparables ne manquent pas et se rencontrent dans des groupes de parasites très variés.
Les processus de favorisation doivent être interprétés comme des adaptations parmi d’autres, sélectionnées pour leur caractère avantageux quant à la transmission. En ce sens, le parasitisme apporte une illustration très riche à la théorie néo-darwinienne de l’évolution biologique.
D’ailleurs, il ne faut pas oublier que, en habitant un être vivant, le parasite se trouve dans un milieu qui, à grande échelle de temps, évolue. On peut donc poser la question: quelle est l’influence de l’évolution de l’hôte sur l’évolution propre de ses parasites? Par exemple, la cladogenèse des hôtes se traduit-elle par une cladogenèse des parasites? En se basant sur des critères morpho-anatomiques, des arbres généalogiques superposables (du moins en partie) ont été proposés pour certains groupes, tels les monogènes parasites de sélaciens. Si cette évolution parallèle est une réalité, elle soulève une nouvelle question, celle de l’existence de pressions sélectives réciproques de la part de l’hôte et du parasite, définissant les processus connus sous le nom de coévolution . Les analyses génomiques et les calculs des distances génétiques à l’intérieur des groupes d’hôtes et des groupes de parasites permettront, à court ou moyen terme, d’élucider l’interprétation évolutive des systèmes hôte-parasite.
2. Chez les végétaux
Environ 95 p. 100 des plantes parasites sont des Champignons appartenant à tous les grands groupes fongiques. Les 5 p. 100 qui restent sont des Bactéries (Eubacteria et Mycobacteria seulement; cf. BACTÉRIES), des Algues (très peu nombreuses: quelques Dinophycées, Chlorophycées et Rhodophycées), une Gymnosperme (Podocarpus ustus , de Nouvelle-Calédonie), des Angiospermes dicotylédones (certaines familles seulement: cf. ANGIOSPERMES). Aucune Bryophyte, aucune Ptéridophyte (lycopodes, prêles, fougères), aucune Angiosperme monocotylédone n’est parasite.
Les hôtes des parasites végétaux sont surtout des plantes vasculaires et, presque aussi nombreux, des animaux terrestres: Insectes (tous les grands groupes; cf. INSECTES), Mammifères (principalement domestiques), homme. On connaît aussi des Champignons parasites d’autres Champignons, ainsi que des Algues parasites d’Algues. Les Dinophycées parasites, organismes exceptionnels, vivent dans des Copépodes marins [cf. CRUSTACÉS].
Du saprophytisme au parasitisme
Bactéries et Champignons présentent, en commun, le caractère d’être aplastidiés (leurs cellules ne contiennent pas de plastes, organites intracellulaires dont la vocation est de produire de la chlorophylle) et sont fondamentalement hétérotrophes, c’est-à-dire incapables d’élaborer, à partir d’éléments minéraux, les matériaux organiques (nutrilites et vitamines) nécessaires à la construction et à la multiplication de leurs cellules. Nombreux sont ceux qui prélèvent ces matériaux dans les milieux organiques naturels (corps en décomposition, humus): on les appelle saprophytes . Or beaucoup de saprophytes manifestent une propension plus ou moins marquée à rechercher leur nourriture organique chez un hôte vivant, en d’autres termes à devenir parasites. Tous les degrés existent dans cette propension: le saprophyte ne devient parasite que dans des conditions plus ou moins rarement réalisées; ou bien, au contraire, le parasitisme est la règle et le retour à la vie saprophytique exceptionnel ou impossible. Tous les intermédiaires existent ainsi, chez les Bactéries et les Champignons, entre les parasites facultatifs, les parasites préférentiels et les parasites obligatoires.
Effets toxiques et toxi-infectieux
De nombreux Champignons et Bactéries pathogènes sont, en réalité, des saprophytes: ayant pénétré dans l’hôte par ses orifices naturels ou par des blessures, ils produisent une toxine diffusible. Chez les animaux, cette exotoxine se répand très vite par voie sanguine, entraînant rapidement la mort (diphtérie, tétanos). Chez les végétaux, sans milieu intérieur, la diffusion lente crée, autour de l’agresseur (nombreux «parasites de blessures»), une auréole de cellules mortes, dans laquelle il pénètre et qu’il étend à mesure qu’il se développe, causant des destructions finalement très étendues. Dans tous les cas, la Bactérie ou le Champignon ne prolifèrent que dans des tissus préalablement tués (parasites nécrotrophes ).
Plus généralement, la toxine produite par le Champignon ou la Bactérie est peu diffusible (endotoxine): la destruction des tissus de l’hôte et la prolifération du microbe sont concomitantes. Ces processus peuvent être rapides (infections aiguës, qui sont propres à l’organisme animal, où le sang facilite grandement la propagation de l’agresseur) ou lente (infections chroniques: abcès «froids» de la tuberculose humaine; chancres des végétaux).
Effets tuméfiants
Les tumeurs produites chez des animaux ou des végétaux, par certains Champignons ou Bactéries parasites, croissent lentement et n’atteignent qu’un volume limité; elles n’offrent aucun caractère cancéreux. Tels sont les mycétomes de l’homme et des animaux domestiques, causés par certains Actinomycètes et Champignons parasites, ainsi que les galles bactériennes observées chez des végétaux divers. Une mention spéciale doit être faite d’Agrobacterium tumefaciens , agent des galles du collet (crown gall ) de nombreux végétaux. Cette Bactérie provoque des tumeurs, dans lesquelles les cellules ont subi une modification physiologique profonde: elles continuent de proliférer, même lorsque le microbe a disparu et, greffées sur des hôtes sains, se multiplient indéfiniment en produisant de nouvelles galles. Le microbe induit ainsi, en quelques heures, une transformation durable, qui fait d’une cellule normale, soumise aux corrélations intraorganiques, une cellule proliférant de manière anarchique et illimitée, semblable à une cellule cancéreuse.
Particularités du parasitisme fongique
Les Bactéries parasites préfèrent les animaux, les Champignons parasites les végétaux. Les tissus végétaux, normalement acides, contrairement à ceux des animaux, sont, pour les Bactéries, généralement alcalinophiles, un milieu peu favorable. De plus, les solides parois des cellules végétales offrent, à la pénétration des corps bactériens, des obstacles que ceux-ci ne rencontrent pas dans les tissus animaux. Les Bactéries ne peuvent guère pénétrer dans une plante vasculaire que par des blessures. Tandis qu’un filament mycélien, sorti d’une spore germée sur une feuille, peut s’allonger jusqu’à rencontrer la voie d’entrée facile qu’est un ostiole stomatique. Les hyphes germinatifs de certains Champignons sont même capables de perforer les épaisses cuticules foliaires. À l’intérieur d’une feuille ou d’une tige, les filaments mycéliens, qui forment des ensembles cohérents, s’insinuent dans les lacunes aérifères, dissolvent, par des enzymes pectinolytiques, le ciment qui unit les cellules et désagrège peu à peu les tissus. Certains Champignons (mildious, rouilles, etc.) disposent du remarquable pouvoir de percer les parois cellulaires de l’hôte et d’émettre un diverticule simple ou ramifié (suçoir), qui pénètre à l’intérieur des cellules (fig. 5); ce suçoir y prélève des substances solubles provenant de la désintégration de matériaux appartenant à un protoplasme vivant: cette biotrophie , mécanisme très subtil, est l’inverse de la nécrotrophie (cf. supra , Du saprophytisme au parasitisme ) et unit étroitement le parasite à l’hôte qui lui fournit un aliment spécifiquement élaboré. Dans la biotrophie, qui condamne l’hôte à un affaiblissement progressif allant jusqu’à l’épuisement, l’aspect trophique, privatif, du parasitisme se manifeste souvent à l’état presque pur.
De l’autotrophie au parasitisme
Les faits précédemment relatés ne concernent que des organismes aplastidiés, fondamentalement hétérotrophes: l’évolution du parasitisme à partir du saprophytisme n’est pas surprenante, et l’on comprend que les parasites végétaux appartiennent, en grande majorité, aux Bactéries et aux Champignons. Il est, par contre, étonnant de constater que des organismes chlorophylliens, doués du pouvoir de photosynthèse et donc autotrophes, se sont adaptés à la vie parasitaire. Certains d’entre eux sont restés autotrophes, tout en devenant parasites stricts. Chez d’autres, le pouvoir de photosynthèse s’est affaibli ou a été perdu: mais même dans les cellules de ces parasites sans chlorophylle subsistent, sceau de la photosynthèse ancestrale, des plastes incolores. Ces plantes physiologiquement dégradées par le parasitisme sont aussi morphologiquement transformées: leur appareil végétatif tend à se réduire au système absorbant. Chez tous les autotrophes ou anciens autotrophes devenus parasites stricts, l’aspect organique du parasitisme est évident.
Les Algues parasites
Certaines Chlorophycées sont parasites de plantes vasculaires. Les feuilles persistantes des arbustes du sous-bois, dans les forêts des tropiques humides, sont un substrat favorable pour beaucoup d’organismes: Bactéries, Champignons, Algues vertes, Lichens. La plupart de ces épiphylles , qui se développent lentement sur les cuticules en utilisant les nutrilites excrétés par la feuille, sont des saprophytes. Cependant, certaines Chlorophycées du genre Cephaleuros (Chætophorales), pénétrant, sinon à l’intérieur des feuilles, du moins dans l’épaisseur des cuticules, sont des semi-parasites; C. virescens peut causer, dans certaines conditions, des dégâts appréciables dans des plantations de théiers. D’autres Algues, encore mal étudiées, vivent et accomplissent tout leur cycle dans les parenchymes foliaires; les moins mal connues sont les Phyllosiphon , parasites des Aracées: les filaments siphonés de ces Algues ne contiennent plus de chlorophylle et vivent en parasites stricts; ils forment, en fin de cycle, des spores intensément vertes dont le devenir est inconnu.
Mais c’est chez les Rhodophycées marines que le parasitisme algal est le plus développé et le plus intéressant. Les Algues rouges parasites sont toutes dépourvues de pigments assimilateurs (chlorophylle et phycoérythrine). Deux cas doivent être distingués. Ou bien l’hôte et le parasite sont systématiquement très éloignés; ainsi Harveyella mirabilis (Cryptonémiale) vit aux dépens de Rhodomela confervoides (Ceremiale): le parasite et l’hôte appartiennent à deux ordres différents (alloparasitisme normal). Ou bien, le parasite et l’hôte, bien que distincts (le parasite ne contient pas de chlorophylle et ses caractères morphologiques sont simplifiés par rapport à ceux de l’hôte), paraissent très étroitement et manifestement apparentés: leurs organes reproducteurs sont identiques, et le mode d’insertion du parasite sur l’hôte ne comporte pas de séparation entre les cellules de l’un et de l’autre. Le parasite serait un mutant sans chlorophylle ne pouvant plus vivre en autotrophie et qui se serait fixé sur un individu frère normal (adelphoparasitisme; 見嗀﨎凞﨏礼﨟, frère).
Les végétaux vasculaires parasites
Les végétaux vasculaires parasites appartiennent à certaines familles seulement et sont tous parasites obligatoires.
La plupart des Scrophulariacées (Tubiflorales) parasites sont annuelles (Rhinanthus , Euphrasia , Striga , etc.); produisant en grand nombre des graines minuscules, elles disposent d’un pouvoir d’infestation élevé, renouvelé chaque année. Dans le sol, leurs racines émettent des suçoirs dans celles des espèces voisines. Leur tige feuillée est verte, et le parasite ne prélève, dans l’hôte, que de la sève brute (hémiparasitisme ), ce qui peut suffire, cependant, à l’affaiblir considérablement.
Les cuscutes (Convolvulacées, Tubiflorales), elles aussi annuelles et produisant d’innombrables petites graines, n’ont plus de feuilles, et leur pouvoir photosynthétique est diminué (teneur en chlorophylle peu élevée). Leurs tiges, filiformes et volubiles, enlacent les rameaux feuillés de leurs hôtes: à chaque point de contact, la cuscute émet un suçoir qui pénètre jusqu’aux tissus conducteurs de l’hôte.
D’autres végétaux vasculaires sont pérennes, et leurs hôtes sont aussi des plantes pérennes, généralement des arbustes ou des arbres.
Les Loranthacées (gui, par exemple; ordre des Santalales) n’ont plus de racines; elles vivent fixées sur les branches des arbres dans lesquelles elles prélèvent toute la sève brute qui leur est nécessaire. Mais leurs feuilles sont vertes et normalement développées. Ces plantes sont parasites parce que dépourvues de racines et non parce que leur pouvoir de photosynthèse est déficient.
Les Orobanchacées (Tubiflorales), Balanophoracées (Santalales) et Rafflésiacées (Aristolochiales) n’élaborent plus de chlorophylle. Ces plantes, physiologiquement très dégradées, dépendant de leur hôte pour toute leur nutrition minérale et organique, sont aussi fortement marquées morphologiquement. Leur appareil végétatif est très réduit: chez Thonningia , Balanophoracée euryxène parasite des arbres de la forêt tropicale africaine, cet appareil consiste en racines souterraines, ou cordons (fig. 2), qui captent celles des hôtes; en pénétrant dans le parasite, chaque racine détermine la formation d’une galle dans laquelle les faisceaux libéro-ligneux de l’hôte se ramifient en d’innombrables petites branches; ces ramifications assurent l’irrigation, donc la nutrition des tissus du parasite. Chez les Rafflésiacées, l’appareil végétatif consiste en cordons parenchymateux anastomosés, sans vaisseaux, s’insinuant, comme un mycélium de Champignon, dans le cambium de l’hôte. Dans les trois familles, l’appareil végétatif produit, à l’extérieur de l’hôte, des inflorescences ou des fleurs solitaires. Les fleurs (cf. ANGIOSPERMES, planche II: fleurs de clandestine; FLEUR, photo d’une fleur de Rafflesia , la plus grande fleur connue, dont le diamètre atteint un mètre) s’épanouissent au niveau du sol ou au sommet d’une courte hampe florale; celles des Balanophoracées, minuscules, sont groupées sur une inflorescence ressemblant à un carpophore de Champignon.
parasitisme [ parazitism ] n. m.
• 1719; de parasite
1 ♦ Condition d'un être vivant qui vit sur un autre en parasite (II).
2 ♦ Présence de parasites dans un organisme, dans un organe. Parasitisme intestinal.
3 ♦ Mode de vie du parasite (I). « Avec l'ignoble tendance au parasitisme qu'avaient les gens de sa génération » (Montherlant).
● parasitisme nom masculin Présence et mode d'existence des parasites dans certains organes ou tissus. État d'un organisme qui vit aux dépens d'un organisme d'une autre espèce, que l'on appelle l'hôte. État, mode de vie de quelqu'un qui vit aux crochets d'autrui : Parasitisme social.
parasitisme
n. m.
d1./d état du parasite, de la personne qui vit aux dépens d'autrui.
d2./d BIOL Condition de vie d'un parasite.
⇒PARASITISME, subst. masc.
A. —BIOL. [Corresp. à parasite I B 1; p.oppos. à commensalisme (dér. s.v. commensal) et à prédatisme (rem. s.v. prédation)]
1. (Condition de) vie d'un parasite (animal ou végétal), d'un être vivant en général qui subsiste aux dépens d'un autre. Parasitisme des Protozoaires; aspects divers du parasitisme; évolution du parasitisme. Il est un ordre de faits (...) qui ont été particulièrement mis en évidence à la fin du XIXe siècle, ce sont ceux relatifs au parasitisme, c'est-à-dire la vie obligatoire de certaines espèces sur —ou dans —d'autres espèces rigoureusement définies pour chaque cas, et aux dépens de celles-ci (Hist. gén. sc., t.3, vol. 1, 1961, p.414). Le bilan général du parasitisme se solde par une perte pour l'exploité et un gain pour l'exploitant. Il semble donc, à première vue, assez facile de séparer le parasitisme du commensalisme (Biol. t.2 1970):
• ♦ Le parasitisme, entraîne donc, de par la spoliation de l'hôte, un état plus ou moins morbide qui définit la maladie parasitaire (...). Les modalités du parasitisme sont multiples selon que les parasites sont fixes ou errants. Ainsi on peut reconnaître un parasitisme intermittent comme chez les Diptères adultes: mouches, moustiques...; un parasitisme continu comme chez les Acariens: gales, poux, etc.
Animaux 1981.
— En partic. État de parasite; mode d'existence des parasites. Parasitisme continu ou provisoire. Le parasitisme de l'albumen (...) se marque parfois par la différenciation de véritables suçoirs (PLANTEFOL, Bot. et biol. végét., t.1, 1931, p.51).
♦Parasitisme absolu. ,,Propriété d'un virus de ne pouvoir se reproduire qu'à l'aide des ARN messagers et des ribosomes de sa cellule-hôte. C'est l'un des cinq caractères fondamentaux qui définissent un virus`` (Méd. Biol. t.3 1972).
2. Présence de parasites chez un être vivant (dans son sang, dans l'un de ses organes). Parasitisme sanguin, intestinal. Il est facile d'éliminer (...) les pigmentations localisées d'origine externe, telles que celles qui succèdent (...) au parasitisme cutané (JOSUÉ, GODLWESKI ds Nouv. Traité Méd. fasc. 8, 1925, p.354).
3. État d'un organisme parasité, infesté par des parasites. En effet, ces maladies se montrent sous forme de cas isolés, d'apparence accidentelle, comme s'il s'agissait du parasitisme fortuit chez l'homme de champignons répandus dans la nature (LONGERON ds Nouv. Traité Méd. fasc. 4, 1925, p.478).
B. —[Corresp. à parasite I A 3; à propos d'une pers. à l'égard d'une autre pers. ou de la collectivité]
1. Mode d'existence d'un individu qui (à l'instar de l'animal parasite) vit et prospère aux dépens d'autrui. La sécurité de mon parasitisme est compromise. J'ai dispersé les plumes de mon nid douillet. Il va falloir déguerpir (RENARD, Écorn., 1892, p.115). On lit d'avance, dans le regard des fortunes (...) le soupçon (...) de l'adresse obligée, d'une espèce de parasitisme, d'écorniflage qu'ils tolèrent (ARNOUX, Solde, 1958, p.23).
2. Condition, état (de l'individu qui vit ainsi en) parasite. Il [l'impôt] attaque le travailleur dans sa liberté et dans sa conscience, dans son corps et dans son âme, par le parasitisme, les vexations, les fraudes qu'il suggère, et la pénalité qui les suit (PROUDHON, Syst. contrad. écon., t.1, 1846, p.294). De temps en temps seulement (...) elle remettait et faisait redescendre Anatole à l'humble place qu'il avait dans la maison, à l'infériorité et au parasitisme de sa position (GONCOURT, Man. Salomon, 1867, p.338). Les ouvriers reprochent leur parasitisme à ceux qu'ils nomment ronds de cuir (WILBOIS, Comment fonct. entr., 1941, p.12).
3. En partic. [Au niveau soc.] Parasitisme social. Le parasitisme consommant improductivement une partie de la richesse collective, il se fait dans le produit net du travailleur un vide qui restreint ses moyens d'échange (PROUDHON, Créat. ordre, 1843, p.278). Il faut réagir contre la tendance de certains sociologues à généraliser la notion de parasitisme pour l'introduire en sociologie; tout être qui vit et prospère aux dépens d'autrui serait pour eux un parasite; les sinécures, la simple concurrence économique, seraient des faits de parasitisme (GOBLOT 1920). Le parasitisme social peut prendre des formes très variées, depuis la délinquance, l'e.ploitation, le chantage, la corruption, la violence pour s'assurer des moyens ordinaires d'existence jusqu'à la prostitution et la mendicité (BIROU 1966).
4. Au fig. [Corresp. à parasite II; à propos d'un inanimé] Caractère encombrant, perturbateur ou nuisible d'une chose et p.méton. cette chose considérée comme une tare. La crise elle-même et les questions qu'elle soulève en moi seront la matière des derniers fragments d'une étude que je débarrasserai du parasitisme des souvenirs insignifiants (BOURGET, Disciple, 1889, p.65). L'idéalisme de la vie intérieure, qui méprise l'argent, comme un parasitisme, oublie qu'il n'en a le droit que s'il y a renoncé; mais l'idéalisme qui pousse sur un terreau argenté et prétend s'en désintéresser, est le pire parasitisme (ROLLAND, Âme ench., t.2, 1925, p.55).
Prononc. et Orth.:[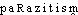 ]. Att. ds Ac. dep. 1878. Étymol. et Hist.1. a) 1719 «mode de vie du parasite» (N. GUEUDEVILLE, trad. Les Comédies de Plaute, La Persane, vol. 8, 145, Leyde, Vander Aa ds QUEM. DDL t.7); b) 1808 «fait de vivre aux dépens de la société» (Ch. FOURIER, Théorie des quatre mouvements, IIIe part., 6, concl. ds DUB. Pol.: le «Parasitisme» [...] spolie le Corps social); 1898 parasitisme social (MASSART et VAN DER VELDE, Parasitisme organique et parasitisme social, Paris ds Gde Encyclop.); 2. 1832 biol. (A. G. DE CANDOLLE, Physiol. végét., I, 1462 ds Fonds BARBIER: De l'influence que les végétaux exercent les uns sur les autres [...] Du faux parasitisme); 1899 parasitisme vrai (Gde Encyclop.); 3. 1966 psychiatrie parasitisme mimique (MOOR). Dér. de parasite; suff. -isme. Cf. angl. parasitisme, att. en 1611 ds COTGR., s.v. escorniflerie. Fréq. abs. littér.:37.
]. Att. ds Ac. dep. 1878. Étymol. et Hist.1. a) 1719 «mode de vie du parasite» (N. GUEUDEVILLE, trad. Les Comédies de Plaute, La Persane, vol. 8, 145, Leyde, Vander Aa ds QUEM. DDL t.7); b) 1808 «fait de vivre aux dépens de la société» (Ch. FOURIER, Théorie des quatre mouvements, IIIe part., 6, concl. ds DUB. Pol.: le «Parasitisme» [...] spolie le Corps social); 1898 parasitisme social (MASSART et VAN DER VELDE, Parasitisme organique et parasitisme social, Paris ds Gde Encyclop.); 2. 1832 biol. (A. G. DE CANDOLLE, Physiol. végét., I, 1462 ds Fonds BARBIER: De l'influence que les végétaux exercent les uns sur les autres [...] Du faux parasitisme); 1899 parasitisme vrai (Gde Encyclop.); 3. 1966 psychiatrie parasitisme mimique (MOOR). Dér. de parasite; suff. -isme. Cf. angl. parasitisme, att. en 1611 ds COTGR., s.v. escorniflerie. Fréq. abs. littér.:37.
]. Att. ds Ac. dep. 1878. Étymol. et Hist.1. a) 1719 «mode de vie du parasite» (N. GUEUDEVILLE, trad. Les Comédies de Plaute, La Persane, vol. 8, 145, Leyde, Vander Aa ds QUEM. DDL t.7); b) 1808 «fait de vivre aux dépens de la société» (Ch. FOURIER, Théorie des quatre mouvements, IIIe part., 6, concl. ds DUB. Pol.: le «Parasitisme» [...] spolie le Corps social); 1898 parasitisme social (MASSART et VAN DER VELDE, Parasitisme organique et parasitisme social, Paris ds Gde Encyclop.); 2. 1832 biol. (A. G. DE CANDOLLE, Physiol. végét., I, 1462 ds Fonds BARBIER: De l'influence que les végétaux exercent les uns sur les autres [...] Du faux parasitisme); 1899 parasitisme vrai (Gde Encyclop.); 3. 1966 psychiatrie parasitisme mimique (MOOR). Dér. de parasite; suff. -isme. Cf. angl. parasitisme, att. en 1611 ds COTGR., s.v. escorniflerie. Fréq. abs. littér.:37.
parasitisme [paʀazitism] n. m.
ÉTYM. 1719; de parasite, I. et II., et -isme.
❖
———
1 Mais la Patience est forte dans le Parasitisme : c'est sa vertu dominante, régnante toujours triomphante.
Plaute, la Persane, trad. 1719, in D. D. L., II, 7.
2 Avec l'ignoble tendance au parasitisme qu'avaient les gens de sa génération, mâles et femelles, durant ces années-là, elle vivait six mois en Bretagne, chez la cousine à héritage, et six mois à Paris, chez des amis (…)
Montherlant, les Célibataires, II, I.
———
II (1832).
1 Biol., cour. État d'un être vivant qui vit sur un autre (parasitisme vrai, commensalisme, mutualisme…). On dit aussi, dans ce sens, parasitisme faux ou pseudo-parasitisme. — Biol. Mode de vie du parasite (II., 1.), dit parfois parasitisme vrai (→ Parasite, cit. 8). || Parasitisme intraspécifique, où le mâle, profondément dégradé, est parasite de la femelle.
2 Présence de parasites dans un organisme, dans un organe. || Parasitisme intestinal.
❖
COMP. Pseudo-parasitisme, xénoparasitisme.
Encyclopédie Universelle. 2012.